
A vírusokat mindig is az élő és az élettelen határterületére helyezte a tudomány. Határozott bekategorizálás mindezidáig nem történt, már csak azért sem, mert magának az életnek sincs jól körülírt, pontos definíciója. Érvek és ellenérvek sokasága hangzott már el a vírusok élő mivoltával kapcsolatban, de ezek egyelőre úgy tűnik, kioltják egymást, hiszen határozott álláspont továbbra sincs.
Az egyik, legtöbb vitára okot adó tényező az óriásvírusok kérdése, melyek hatalmas, baktériumokéval vetekedő genomméretük miatt jelentős fejtörésre késztetik a virológusokat és genetikusokat. Egyesek szerint az élet eddig ismeretlen, negyedik doménjéből származnak, mások azonban elvetik ezt a feltevést és pusztán csak különleges evolúciós illetve adaptációs jelenségnek tartják létüket. Mára gyakorlatilag új kutatási terület szerveződött az óriásvírusok megismerése köré.
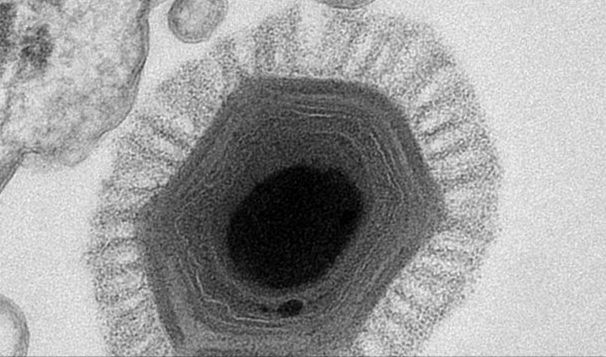
A vírusok mint biológiai entitások már a huszadik század elejétől ismertek. Gyakorlatilag nem mások, mint fehérjeburokkal (kapsziddal) körbevett DNS- vagy RNS-darabok. A kapszid alakja miatt nagy morfológiai változatosságot mutatnak (például helikális, ikozaéderes alak). Egyetlen „élet”funkciójuk a szaporodás, önmaguk lemásolása. A vírusok az élővilág összes doménjében képesek sejteket fertőzni, eukariótákat, baktériumokat és archeákat egyaránt. Szaporodásukhoz azonban feltétlenül szükségük van gazdasejtre. „Életciklusuk” meglehetősen egyszerű: a specifikus kapszid-sejtfelszíni receptor kapcsolat segítségével megtapadnak a sejt felszínén, majd a DNS-t vagy RNS-t bejuttatják a gazdasejtbe. Az örökítőanyag ezután beépül a gazdasejt genomjába és elkezdi magát több példányban lemásolni. Végül az új vírusok összeszerelődnek, majd elhagyják a sejtet, gyakran annak pusztulását okozva. A vírusok sejten kívüli alakját virionnak nevezik, melyek átlagos mérete néhány tíz és néhány száz nanométer között mozog. A kapszid fizikai méreténél azonban informatívabb adat a vírus genommérete, amely minimum két kilobázis nagyságú és legalább négy gént kell tartalmaznia.
A vírusok méretének felső határa azonban jóval magasabb, mint amilyennek gondolnánk. A mindössze négy éve leírt pandoravírusok fizikai mérete meghaladhatja az 1 mikrométert, genomméretük pedig a két és fél megabázist, több mint 2500 fehérjekódoló génnel. Összehasonlításképp egy Escherichia coli sejt alig kétszer ekkora genommal és fizikai mérettel bír. Mindezek tehát figyelemreméltó mutatók egy olyan entitástól, amelyet sokan nem is tartanak élőnek és egyetlen tevékenysége más sejtek megfertőzése.

A pandoravírusokon túlmenően számos egyéb óriásvírust is leírtak. A kilencvenes évek végén találtak rá az első mimivírusokra, melyek amőbákat használtak gazdasejtként. A legelső felfedezett óriásvírus az Acanthamoeba polyphaga mimivírus volt, melyet érdekes festődési tulajdonságai – és mérete – alapján először baktériumnak hittek (az már korábban ismert volt, hogy léteznek olyan baktériumok, amelyek a vírusokhoz hasonlóan csak gazdasejt segítségével maradnak életben). A neve is innen származik: a baktériumok kinézetének „mimikálásáért” kapta. Ezt követte a megavírusok leírása, melyek szintén amőbákat fertőznek, de a mimivírusoknál pár százzal több génjük van. Ennek megfelelően a fertőzési idejük is hosszabb, mintegy öt órával több időbe telik. Ezután következtek a már említett pandoravírusok. Ekkorra már meglehetősen nagy „kultuszuk” alakult az óriásvírusoknak a tudósok körében. Rengeteg virológus, genetikus, bakteriológus és taxonómus tűzte ki célul megismerésüket, genomjuk és az általuk kódolt fehérjék felderítését, de leginkább rokonsági kapcsolataiknak feltárását. Erre pedig egyre bővülő lehetőségeik lettek a XXI. század előrehaladtával, hiszen a molekuláris biológia eszköztára soha nem látott fejlődésnek indult. A vírusok, így az óriásvírusok sem képeznek fosszíliákat, genomszekvenciájukat azonban minden további nélkül meg lehet állapítani és annak ellenére, hogy nem tekinthetők egyértelműen élőknek, azonosítani lehet az általuk kódolt fehérjéket és rokonsági kapcsolataikat, a sejtes élőlényekhez hasonlóan pusztán a genomjuk elemzéséből következtetni lehet az evolúciós, filogenetikai viszonyaikra. Kiderült, hogy a legtöbb óriásvírus meglehetősen közeli rokon, genomjukban sok gén nagymértékben konzervált, azaz több csoportjukban is előfordul, viszonylag változatlan formában. A mimivírusoknak például több mint 20 génjük fordul elő nagy számban a Nucleocytoplasmic Large DNA Viruses (NCLDV) gyűjtőnéven emlegetett csoportban.
A legérdekesebb jelenség azonban az, hogy az óriásvírus genomok hajlamosak aminoacil-tRNS szintetázokat, eukarióta iniciációs és elongációs faktorokat, valamint számos egyéb transzlációhoz szükséges fehérjét kódolni. A transzláció a gének fehérjékben történő kifejeződésének utolsó lépése, mely során a DNS-ről készült RNS-átiratból végül fehérje lesz. Ez a legtöbb energiát igénylő és legösszetettebb anyagcserefolyamat a sejten belül, amire elméletben csak sejtes szervezetek képesek. Habár az óriásvírusok között még nem ismert olyan, amely a teljes fehérjeszintetizáló apparátust kódolná (például riboszomális fehérje-géneket még nem találtak bennük), a fehérjéik szintetizálásához szükséges faktorok egy részét azonban már ők maguk biztosítják, ahelyett, hogy a gazdasejt gépezetét használnák, ez pedig meglehetősén „élő” tulajdonság.
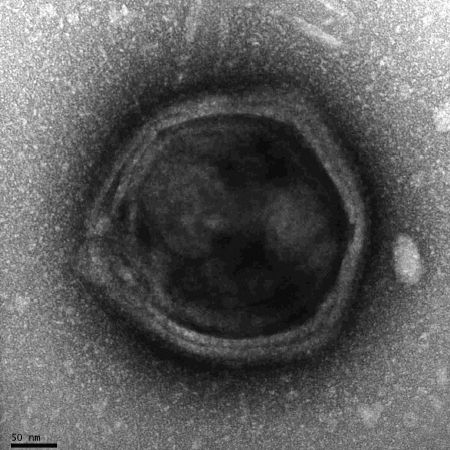
A legtöbb fehérjeszintetizáló faktort a nemrégiben megismert klosneuvírusok kódolják. A klosneuvírusokat egy amerikai-osztrák kutatócsoport fedezte fel az ausztriai Klosterneuburg szennyvíztisztítójában, merő véletlenségből, miközben a nitrifikáló baktériumokat vizsgálták. A genomjuk szekvenálása után pedig igencsak meglepő eredményeket kaptak. A korábban már megismert óriásvírus-genomok alapján számítani lehetett arra, hogy az új csoport is tartalmazni fog a transzlációs apparátus fehérjéinek kifejezéséhez szükséges géneket, valamint további sejtes, sőt, eukarióta szervezetekre jellemző géneket, a klosneuvírusok „felszereltsége” mégis váratlanul érte a kutatókat. 25 féle tRNS szintézisére képesek, melyek legalább 14 féle aminosavhoz tartozó antikodont tartalmaznak. Ezen túlmenően további negyven transzlációhoz köthető fehérjét kódolnak, köztük tizenegy eukarióta transzláció inifiációs és elongációs faktorral, valamint 19 aminoacil-tRNS szintetázzal. Összehasonlításképp a mimivírusok 7 aminoacil-tRNS szintetázt kódolnak.
Ez a rendkívül gazdag génállomány méltán vetette fel a kérdést, hogy hogyan is jöhetett létre egy – legalábbis a genomjában – ennyire „eukariótaként viselkedő” óriásvírus. Az egyik vélekedés az volt, hogy a klosneuvírusok, illetve az óriásvírusok általában, ősi sejtes szervezetekből egyszerűsödtek le és az élet egy rég letűnt, negyedik doménjét képviselik az eukariótákon, baktériumokon és archeákon túl. A másik vélekedés szerint azonban nincs szó ilyesmiről, inkább az a valószínűbb, hogy az óriásvírusok kisebb vírusokból fejlődtek ki, és az újabb örökítőanyag-fragmenteket egyesével vették fel az eukarióta – jobbára amőba – gazdasejtjeikből, ezért tartalmaznak ennyi eukarióta típusú iniciációs faktort és egyéb transzlációs komponenst.
A kérdés eldöntésére a kutatók átfogó filogenomikai analízist végeztek, melynek eredményeképpen kiderült, hogy a klosneuvírusok transzlációs faktorai nagyfokú rokonságot mutatnak számos eukarióta egysejtű, leginkább egysejtű alga hasonló fehérjéivel. Ez a második elképzelést támasztja alá, mivel a „negyedik domén” hipotézis szerint a klosneuvírusok által kódolt transzlációs faktoroknak egy jól elkülönülő, nagy ágat kellene alkotniuk az eukarióta fehérjék törzsfáján belül, ehelyett meglehetősen közel találhatók a többi eukariótához.
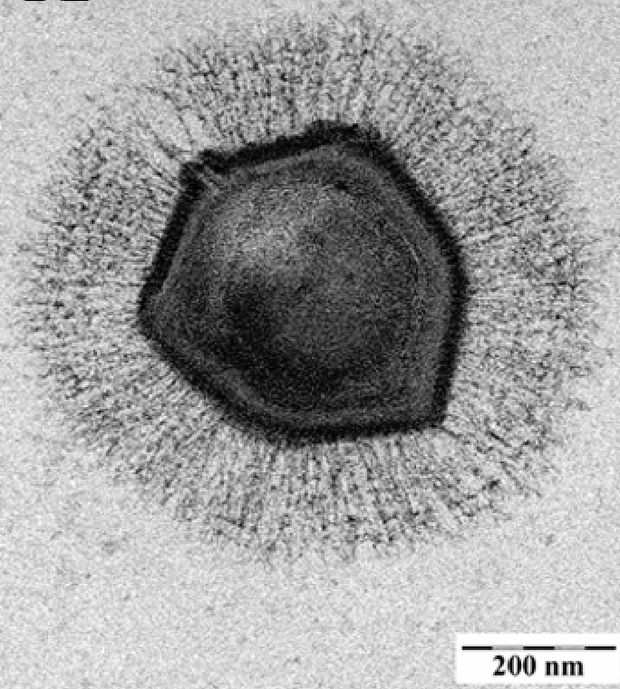
Molekuláris taxonómiai vizsgálatok segítségével igyekeztek meghatározni a klosneuvírusok rokonsági kapcsolatait a többi óriásvírussal is, valamint próbálták azonosítani a lehetséges gazdasejtjeit. Mint kiderült, a klosneuvírusok gazdasejtjeit jórészt a Cercozoa törzs tagjai közt kell keresnünk, ők maguk pedig a mimivírusokhoz állnak legközelebb filogenetikailag, azoknak egy különálló, speciális alcsaládjaként kezelhetők. A kutatók az eredményt öt, az NCLDV csoportban univerzálisan előforduló gén markerként való használatával történő elemzést követően kapták.
Ezt követően elkezdték tüzetesen átvizsgálni a genomjukat, hogy az összehasonlításukból következtetéseket vonhassanak le az óriásvírusok evolúciós dinamikáját illetően. Hét olyan aminoacil-tRNS szintetáz van, amelyet a klosneuvírusok és mimivírusok is kódolnak, ezek közül három monofiletikus, azaz már a közös ősük is tartalmazta őket, mielőtt az evolúciójuk szétvált volna. A maradék négy polifiletikus, tehát már szétválás után, eltérő időpontokban szerezték meg őket. Az elongációs és iniciációs faktorok megoszlása is hasonló kettősséget mutat, utalva arra, hogy az óriásvírusok evolúciója egy szerteágazó és nagyívű folyamat, amely vélhetően az egyes gének szakaszos felvételével, a genom darabról darabra történő kiegészítésével valósul meg.
Ezeken túlmenően az óriásvírusok számos egyéb, fejlett sejtes szervezetre jellemző fehérjét kódolhatnak, így például kollagént illetve nukleinsavak és aminosavak szintéziséhez szükséges enzimrendszereket. Nem találtak még azonban egyetlen olyan óriásvírust sem, amely riboszomális RNS-eket vagy fehérjéket kódolt volna, ami elengedhetetlen lenne az önálló szaporodáshoz és ezen keresztül az élethez. Felmerül azonban a kérdés, hogy ha az óriásvírusok valóban a genomjukhoz történő folyamatos „hozzávarrással” egészítgetik ki fehérjekészletüket és egyre több anyagcsereút enzimrendszerét szerzik be, akkor meddig mehet el a folyamat? Szintén elgondolkodtató, hogy az óriásvírusok génjei között számos olyan található, amely nagymértékű konzerváltságot mutat az összes jelenlegi domént fertőzni képes vírus között, ami arra utal, hogy az óriásvírusok a sejtes életet megelőzően jöttek létre a Földön és kulcsszerepet játszottak az összes további életforma kialakulásában. Annyi bizonyos, hogy hosszú út áll még az óriásvírusok felderítését célzó kutatók előtt, és minden eredmény arra utal, hogy a jövőbeni kutatások még nagyobb és még komplexebb vírusokat tárnak majd fel.
BENKŐ PÉTER
2017/24

